昆明核移植纺锤体胚胎植入
纺锤体的双极化是卵母细胞减数分裂过程中的关键事件之一。近年来,我国学者在人类卵母细胞纺锤体双极化机制研究方面取得了重要进展。通过高分辨成像技术,研究者们揭示了人类卵母细胞纺锤体双极化的独特机制,并发现了调控此过程的关键蛋白。这些研究成果不仅为双折射性纺锤体卵冷冻研究提供了新的视角和思路,也为临床生殖障碍疾病的诊疗提供了科学依据。随着偏光成像技术和冷冻保护剂研究的不断深入,未来有望开发出更加高效、安全的卵母细胞冷冻保存方案。例如,通过改进冷冻速率和程序、优化保护剂配方等手段,进一步减轻冷冻损伤,提高解冻后卵母细胞的存活率和发育潜能。纺锤体的微管在细胞分裂过程中具有自我修复和再生的能力。昆明核移植纺锤体胚胎植入
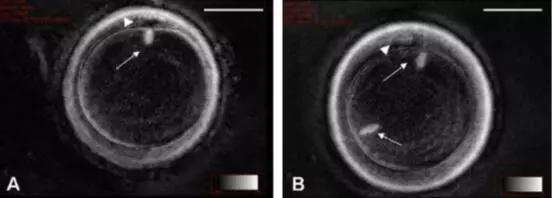
卵母细胞的冷冻保存技术一直是研究的热点之一,特别是针对不同成熟阶段的卵母细胞,如MI期卵母细胞的冷冻保存。MI期卵母细胞具有独特的生物学特性和发育潜能,其纺锤体的稳定性和形态对于后续的受精和胚胎发育至关重要。因此,针对MI期纺锤体卵冷冻的研究不仅具有理论价值,更具有重要的临床应用前景。MI期卵母细胞的纺锤体由微管组成,这些微管结构精细且脆弱,容易受到冷冻过程中温度变化和渗透压变化的影响而发生损伤。纺锤体的损伤不仅会影响卵母细胞的正常发育,还可能导致受精失败或胚胎发育异常。美国Hamilton Thorne纺锤体Hoechst染料纺锤体在细胞分裂中的精确调控是生物体发育的基础。
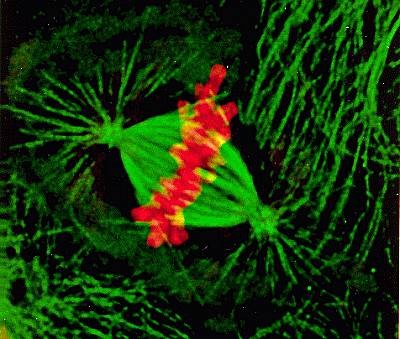
纺锤体的完整性决定了染色体分裂的正确性。在有丝分裂前期,中心体被复制形成两个中心体,并逐渐分离,形成两个纺锤体。纺锤体的微管从中心体发出,与染色体上的着丝粒(kinetochore)结合。着丝粒是一组复杂的蛋白质结构,可以与微管的末端结合。当纤维束的微管末端与着丝粒结合时,纤维束开始缩短,将染色体拉向两端,实现染色体的精确分离。这一过程不仅确保了每个新细胞都能获得正确数量的染色体,还保证了遗传信息的稳定传递。
纺锤体缺陷可以分为多种类型,包括但不限于:微管动力学异常:微管的聚合和解聚速率异常,导致纺锤体结构不稳定。动粒功能障碍:动粒与微管的结合能力下降,影响染色体的正确捕捉和分离。纺锤体检查点失效:纺锤体检查点(spindleassemblycheckpoint,SAC)是确保染色体正确分离的重要机制,其失效会导致染色体分离错误。染色体分离异常:染色体在分裂过程中未能正确分离,导致非整倍体的形成。微管的动态变化是纺锤体功能的关键,任何影响微管聚合和解聚的因素都会导致纺锤体结构的不稳定。例如,某些药物(如紫杉醇)可以稳定微管,但过量使用会导致微管过度稳定,影响纺锤体的正常功能。 纺锤体的形成和功能与细胞的周期调控密切相关。
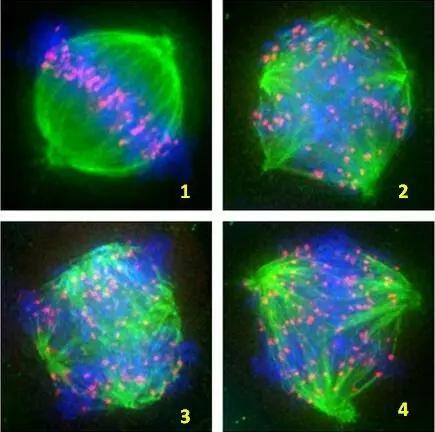
纺锤体生成在含中心体的细胞中,纺锤体的生成开始于细胞分裂前初期-即在细胞核膜分解(NuclearEnvelopeBreakdown,NEB)之前。初期的结构为两个**的以中心体为核的星状体(asters)。当细胞核膜分解后,染色体和星状体发生一系列复杂的互动反应。**终结果为所有的染色体在纺锤体的**(赤道板,)排列整齐,每两条染色体有一个着丝点,每一个着丝点被一束极性相同的微管(通常称为纺锤丝)附着。此时细胞处于分裂中期,纺锤体生成完毕。实验证明,中心体在这个过程中的作用不是必需的。动物细胞在中心体被激光捣毁后仍旧能够筑构纺锤体,但其位置通常不在细胞的大致几何中心,其后的胞质分裂也会受严重影响。纺锤体[1]在不含中心体的细胞中,纺锤体的生成是由染色体本身主导的。此过程由一小分子量的GTP连接蛋白(RanGTPase)控制。细胞核分解后,纺锤丝由染色体周围生成。其后这些纺锤丝会在动力分子与为微管动力的合作影响下自动排列为极性相反大致数目相同的两组。每组的极性相对于一组着丝点。同时在微管远端的动力蛋白dynein会将这些微管束集中到一点,形成纺锤极区(SpindlePolarZone)。与此同时,染色体会自动在赤道板排列整齐。纺锤体生成完毕。纺锤体微管的聚合与解聚受到多种酶的调控。上海无需染色纺锤体卵质量评估
纺锤体的形成需要多种蛋白质的精确协作与调控。昆明核移植纺锤体胚胎植入
光学相干断层成像是一种基于低相干光干涉原理的成像技术,具有高分辨率、非侵入性和实时成像等特点。在纺锤体卵冷冻研究中,OCT技术可用于观察卵母细胞内部结构的细微变化,包括纺锤体的形态和位置。虽然目前OCT技术在纺锤体成像方面的应用还较为有限,但随着技术的不断发展和完善,相信未来OCT将在纺锤体卵冷冻研究中发挥更加重要的作用。虽然MRI和超声波成像在生殖医学中主要用于软组织的成像,如子宫、卵巢等病变检测,但它们在纺锤体卵冷冻研究中的应用也值得探讨。随着技术的不断进步,高分辨率MRI和超声波成像技术可能会实现对卵母细胞内部结构的更精细观察。昆明核移植纺锤体胚胎植入
- 北京鱼精子分析 2025-07-15
- 香港鱼精子分析BCF 2025-07-14
- 上海鱼精子分析W.H.O 6 2025-07-13
- 马精子分析WOB 2025-07-10
- 北京精子分析频闪光源 2025-07-08
- 北京纺锤体实时成像纺锤体改善分级 2025-07-08
- 昆明无需染色纺锤体改善分级 2025-07-08
- 上海纺锤体胚胎植入 2025-07-08
- 深圳非侵入式成像纺锤体胚胎植入 2025-07-08
- 香港偏光成像纺锤体揭示卵母细胞关键结构 2025-07-08
- 徐州步入式老化房 2025-07-21
- 吉林梓盟橡胶加工分析仪要多少钱 2025-07-21
- 山东多语言干式真空泵联系方式 2025-07-21
- 安徽测温装置测温管公司 2025-07-21
- 苏州电子式剪切试验机功率 2025-07-21
- 宁夏保温夹套安全阀供应商 2025-07-21
- 肇庆照明光源检测设备分布光度计欢迎来电 2025-07-21
- 金属检测分析仪检测仪 2025-07-21
- 苏州全自动闪测仪应用 2025-07-21
- 四川高精度称重传感器安装 2025-07-21